

Accumulated Changes In The Genetic Makeup Over Time Results In What?
INTRODUCTION TO POPULATION GENETICS
In this and the next few lectures nosotros will be dealing with population genetics which generally views evolution as changes in the genetic makeup of populations. This is a somewhat reductionist approach: if we could understand the combined action of the forces that change factor frequencies in populations, and and then allow this run over many generations we might understand long term trends in evolution. Standing debate: can the processes of microevolution business relationship for the patterns of macroevolution? Population genetics is an elegant set of mathematical models developed by largely by R. A. Fisher and J. B. S. Haldane in England and Sewall Wright in the The states. Continues to be developed by many mathematical, theoretical and experimental biologists today (encounter J. Crow and M. Kimura Introduction to Population Genetics Theory).
In very simple terms, population genetics involves analyses of the interactions betwixt predictable, "deterministic" evolutionary forces and unpredictable, random, "stochastic" forces. The deterministic forces are oft referred to as "linear pressures" considering they tend to push allele frequencies in one management (up, downwards or towards the middle). Of import forces of this nature are selection, mutation, gene flow, meiotic bulldoze (unequal manual of certain alleles [a grade of selection]), nonrandom mating (also a form of selection). The principal stochastic evolutionary forcefulness is genetic drift which is due to the random sampling of individuals (and genes) in pocket-size populations. It is important to realize that the deterministic forces may act together or confronting 1 another (e.chiliad., choice may "endeavor" to eliminate an allele that is pushed into the population by recurrent mutation). Moreover, deterministic forces may human activity with or against genetic drift, to determine the frequencies of alleles and genotypes in populations (due east.1000., cistron menstruation tends to homogenize different populations while drift tends to make them unlike). Hence, the interaction of these forces is what we are actually interested in (a later lecture), but since this can get very complex mathematically, we will start by analyzing i strength at a time.
To brainstorm nosotros need to sympathize some simple population genetic "accounting." Consider a locus with two alleles (alternative forms of the DNA sequence that "reside" at that locus, e.g., one from mother other from father). Now consider a population of N individuals (Northward=population size); this means that there are 2N alleles in the population. Nosotros can thus talk nearly genotype frequencies and allele frequencies. In a population of N = 100 individuals, if in that location are 25 AA, fifty Aa and 25 aa, then the genotype frequencies are f(AA) = 0.25, f(Aa) = 0.50 and f(aa) = 0.25. If we count up the individual alleles there are 200 of them (because there are 100 diploid individuals). Hence to determine the frequency of the "A" allele we have to count each individual "A" allele that is specified in each diploid genotype. We go f(A) = (25+25+50) / 200 = 0.5. We generally refer to the frequency of the "A" allele as f(A) = p; the frequency of the "a" allele is f(a) = q. Note that p = (1-q) because the sum of the allele frequencies must be one.0. Common "linguistic communication errors" in learning population genetics are to refer to the "p" allele when you actually mean the "A" allele, or to say "the frequency of the p allele" when y'all really hateful: "...p, the frequency of the "A" allele..." Got it?? Skilful.
Since evolution is change in the genetic makeup of a population over fourth dimension, a general approach to modeling this is to determine the allele and genotype frequencies in the adjacent generation (pt+i) that event from the activeness of a force on those frequencies in the current generation (pt). Thus :
pt -> evolution happens -> pt+ane
Consider a simplistic life cycle where the genotypes (a single locus manner of referring to adults) produce gametes. These gametes mate to grade new genotypes (=adults). Run across v.1, pg. 93 and 5.3, pg. 99. The relationship between allele frequencies (sometimes called "factor" frequencies) and genotype frequencies is determined past the Hardy Weinberg Theorem which defines the probabilities by which gametes volition join to produce genotypes. Consider a coin toss: probability of a head = 0.v; of a tail = 0.5; prob. of two heads = 0.5x0.5 = 0.25; prob. of i caput and one tail = 0.5x0.5 = 0.25, etc. Each money is analogous to the blazon of allele you lot tin become from ane of your diploid parents; the tossing of two coins is coordinating to the mating of two individuals to produce four possible genotypes (but heads,tails is the aforementioned as tails,heads). Now consider a scroll of the dice. The probability of each face is 1/6, and is actually coordinating to cases where more than two unlike alleles exist in the population at a given locus. The probability of whatsoever combination is i/6 ten ane/6 = ane/36. But recall that in that location can exist more than than ane way to get many of the combinations (2,three is the same equally 3,2). The general expression for the number of genotypes that can be assembled from n different alleles is: [n(n+1)/2].
Assumptions of Hardy Weinberg: 1) diploid sexual population 2) space size, 3) random mating, 4) no selection, migration or mutation. This is a Null Model; obviously some of these assumptions volition non concur in real biological situations. The theorem is useful for comparing to existent-earth situations where deviations from expectation may point to the action of sure evolutionary forces (e.chiliad., mutation selection, genetic migrate, nonrandom mating, etc.). Use a Punnet foursquare to determine genotype frequencies: f(AA) = p2, f(Aa) = 2pq, f(aa) = q2 and p2 + 2pq + qii = 1 Acquire this: One generation of random mating restores Hardy Weinberg equilibrium. H-Westward equilibrium is when the genotype frequencies are in the proportions expected based on the allele frequencies every bit determined by the relation p2 + 2pq + q2. This is derived more than thoroughly in table 5.ane, and accompanying text, pg. 94.
Example: consider a sample of 100 individuals with the post-obit genotype frequencies:
| Observed Genotype Frequencies | Allele count | Allele frequency | Expected genotytpe frequencies under H-W | |
| BB | 0.71 | 142 B | p = 156/200 = 0.78 | ptwo = (.78)2 = 0.61 |
| Bb | 0.14 | 14 B, xiv b | 2pq = 2(.78)(.22) = 0.34 | |
| bb | 0.15 | xxx b | q = 44/200 = 0.22 | q2 = (.22)2 = 0.05 |
Observed are unlike from expected, thus some forcefulness must be at piece of work to change frequencies.
NATURAL SELECTION
Option occurs because unlike genotypes exhibit differential survivorship and/or reproduction. If we consider a continuously distributed trait (due east.thou., fly length, weight) with a strong genetic basis, the response to selection can exist characterized by where in the distribution the "well-nigh fit" (greatest survivorship&reproduction) individuals lie. If afterwards selection one farthermost is most fit this is directional selection; if the intermediate phenotypes are the nigh fit this is stabilizing pick; if both extremes are the most fit this is disruptive selection.
R. A. Fisher proposed a elementary accounting, or population genetics, approach for one locus with two alleles: nosotros accept AA, Aa and aa in frequencies p2, 2pq, q2 . Ascertain lii as the genotype-specific probability of survivorship, mii every bit the genotype-specific fecundity. We build a model that will predict the frequencies of alleles that will be put into the gamete pool given some starting frequencies at the preceding zygote stage;
| Genotypes | Zygote | -----> -----> | Developed | -----> -----> | Gametes |
| AA | p2 | lAA p2 | thouAA lAA pii | ||
| Aa | 2pq | lAa 2pq | mAa lAa 2pq | ||
| aa | q2 | 50aa q2 | maa fiftyaa q2 |
The gamete column is what determines the frequencies of A and a that will be put into the gamete pool for mating to build the side by side generation'south genotypes. We can simplify by referring to the fitness of a genotype as wtwo = mtwo 50ii . These fitness values will determine the contribution of that genotype to the side by side generation. Thus the frequency of A allele in the next generation pt+1 (sometimes referred to as p') would exist the contributions from those genotypes conveying the A allele divided by all alleles contributed by all genotypes:
pt+1 = (wAA p2 + wAa pq)/(wAA p2 + wAa 2pq + waa q2). Or for the a allele,
qt+ane = (waa qii + wAa pq)/(wAA pii + westAa 2pq + waa q2). Notation that the heterozygotes are not 2pq just pq because in each case they are just being considered for the one allele in question. If we scale all wii's such that the largest = 1.0 we refer to these as the relative fitnesses of the genotypes. A worked example where p = .4, q = .6 and wAA = 1.0 westwardAa = 0.8 westaa = 0.6:
Genotype frequencies are pii = 0.16, 2pq = 0.48, q2 =0.36, thus:
pt+1 = ((.sixteen ten one.0) + (.24 x .8))/((.xvi x ane.0) + (.48 x .viii) + (.36 ten .6)) = .463; and so q = .537 and thus f(AA)t+1 = .215, f(Aa)t+1 = .497 and f(aa)t+1 = .288. Note both allele frequencies and genotype frequencies have changed (compare to what we saw with inbreeding). This tin be continued with the new allele frequencies then on. When volition the selection procedure stop? when D p = 0, i.e., when pt+1 = pt . In some situations this volition terminate but when one allele is selected out of the population (p = 1.0).
Now we tin can consider diverse regimes of selection (southward = selection coefficient, (i-s) is fitness):
| AA | Aa | aa | ||
| I | i | ane | 1 - s | selection against recessive |
| Ii | 1 - s | 1 - due south | 1 | selection against ascendant |
| Iii | 1 | ane - hs | 1 - s | incomplete authority (0<h<1) |
| Four | 1 - s | i | 1 - t | selection for heterozygotes |
Substitute the fitnesses (due westii) in status I to a higher place into the expression D p = pt+i - pt and prove for yourself that the equations on folio 101 (eqn. 5.v) is related to the expression for pt+ane shown higher up. Get-go three are directional in that selection stops merely when allele is eliminated. In I the elimination process slows down considering every bit q becomes small the a alleles are ordinarily in heterozygote country and in that location is no phenotypic variance. In Ii choice is slow at commencement considering with q small most genotypes are AA and then there is low phenotypic variance; as option eliminates A alleles q increases and the frequency of the favored genotype (aa) increases so selection accelerates. 3 is like the worked example run to fixation/loss. IV is known every bit balancing option due to overdominance (heterozygotes are "more" than either homozygote). Both alleles maintained in population past pick. This is an example of a polymorphic equilibrium (fixation/loss is also an equilibrium status only information technology is not polymorphic). The frequencies of the alleles at equilibrium will be:
pequil = t/(southward + t); qequil = s/(s+t).
Classic instance = sickle cell anemia. A=normal allele; South=sickle allele. S should exist eliminated because sickle cell anemia lowers fettle. S is maintained where malarial agent (Plasmodium falciparum) exists because AS heterozygotes are resistant to malaria. Note that S allele is very low frequency where there is no malaria (the selective coefficient of S is dissimilar because the environment is dissimilar). See figure 5.8, pg. 120; table 5.ix, pg. 119.
Another way that genetic variation tin be maintained is through multiple niche polymorphism (polymorphism maintained past environmental heterogeneity in selection coefficients). If different genotypes are favored in different niches, patches or habitats, both alleles can be maintained.
| AA | Aa | aa | |
| habitat 1 | ane.0 | 0.eight | 0.5 |
| habitat two | 0.5 | 0.8 | 1.0 |
Heterozygotes will accept the highest boilerplate fitness although they are non the most fit in either habitat (run across figure v.12, pg. 124). The same dynamics would apply to temporal heterogeneity (spring and fall; winter and summer) assuming that pick did not eliminate one allele during the first period of selection. Archetype example of temporal heterogeneity: tertiary chromosome inversions of Drosophila pseudoobscura studied by T. Dobzhansky. Different chromosomal arrangements ("Standard" and "Chiricahua") show reciprocal frequency changes during the year.
Yet some other fashion to maintain variation by selection is through frequency dependent selection.
If an allele'due south fettle is non constant but increases as it gets rare this will drive the allele back to higher frequency. See effigy 5.nine, pg. 121. Example: allele may give a new or distinct phenotype that predators ignore because they search for food using a "search image" (e.g., I like the green ones).
Almost (by no ways all) evolutionary biologists believe that selection plays a major role in shaping organic diverseness, but it is often difficult to "see" selection. Ane reason is that selection coefficients tin exist quite small-scale (1-s ~1) so the response to choice is small. When choice coefficients are large D p tin be large, simply the problem here is that with directional selection fixation is reached in a few generations and we yet can't "see" selection unless we are lucky enough to catch a population in the middle of the period of rapid change.
What affects the rate of change under selection? Recall that D p = pt+i - pt
D p = [(wAA p2 + wAa pq)/(wAA p2 + wAa 2pq + waa q2)] - p . With some simple algebra we tin can rearrange this
equation to: D p = (pq[p(wAA - wAa) + q(westwardAa - westaa)])/(wAA p2 + due westAa 2pq + waa q2)
Note that D p will be proportional to the value of pq. This value (pq) will exist largest when p=q=0.5 or, in English language, when the variance in allele frequency is greatest. This is a simplified version of the primary point of the fundamental theorem of natural selection modestly presented by R. A. Fisher.
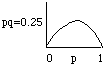
It states that the rate of evolution is proportional to the genetic variance of the population. In the above example we have non explicitly defined the fitnesses wiis or the authorization relationships and these tin take a major issue on D p equally written above.
Some other important observation for looking at this D p equation and plugging in some values is that selection ever increases the hateful fitness of the population. For example with p=0.four, q=0.6 and wAA=1, wAa=0.8 and waa=0.half-dozen, the mean fitness (due west'bar') = 0.76. After one generation of selection p' = 0.463 and q' = 0.537. Recalculating w'bar' we go wbart+ane = 0.78, which is greater than 0.76. When will this procedure cease? At fixation (or equilibrium with overdominance).
This treatment of the algebra of natural selection illustrates what option lone can do to allele and genotype frequencies. In the side by side lectures we will consider other evolutionary forces (mutation factor menses, genetic drift), how they act alone, and somewhen, how they interact with each of the other evolutionary forces.
Source: https://biomed.brown.edu/Courses/BIO48/6.PopGen1.HW.drift.HTML
Posted by: deanegrapinglies.blogspot.com
0 Response to "Accumulated Changes In The Genetic Makeup Over Time Results In What?"
Post a Comment